精读笔记
Problem Setting
[Radial Interaction Tomography: Recognizing Non-Transitive Evolutionary Games from One Range-Expansion Image](arXiv preprint / 2026)。这篇论文实际解决的是一个 endpoint inverse problem:从单张径向扩张终态图像中恢复可见 genotype 接触边界上的 radius-indexed pairwise flow,并判断这些 flow 是否能由某个标量 fitness hierarchy 解释。真正困难点不是视觉分割,而是可辨识性:终态图像确实保存了某种历史,但只保存为径向 deposition coordinate 下的几何轨迹,不自动等价于物理时间、孤立扩张速度或机制参数。以前方法主要把 sector geometry 用来估计常数选择优势或边界随机漂移,默认竞争近似可标量化;一旦存在 cyclic dominance 或局部 antagonism,这类标量解释会把非势结构压扁。关键矛盾是:单图像信息很少,但径向冻结又使它不是普通静态图像;论文要做的是最大化利用这点,同时不越界解释。
Motivation
已有路线不够,是因为它们把 colony pattern 当作 selection/fitness 的间接读数,而不是先问 endpoint geometry 本身到底识别了什么。作者的核心观察是,sector boundary 在 log-polar 坐标中形成一组曲线,曲线角度对半径坐标的导数天然是定义在接触图边上的有向 flow;如果竞争真能由 scalar hierarchy 表示,这些 edge flows 必须是节点势的梯度。缺口在于:非传递竞争并不缺生物例子,缺的是从单张终态图像中识别非传递 signature 的可证伪几何判据。换言之,论文动机不是引入新生态机制,而是防止把所有 sector curvature 都粗暴解释为 fitness,并给出一个 endpoint-only 可支持的最强声明。
Core Idea
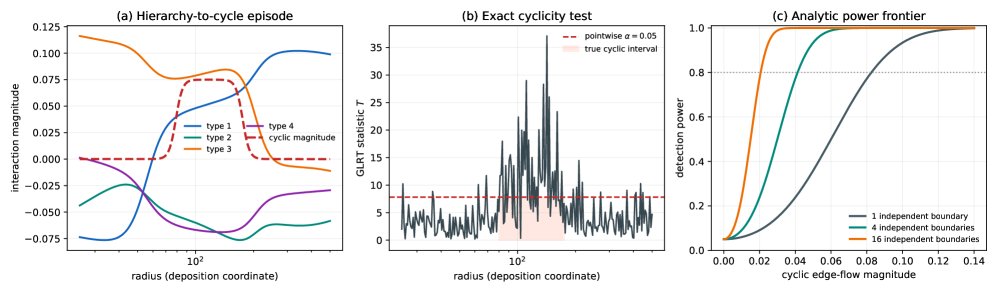
核心思想是把图像中的边界曲线重写成接触图上的 edge-flow field。对每条 i-j 边界,提取 θ_ij(ρ),再用 q_ij(ρ)=dθ_ij/dρ 表示局部角向侵占速度;随后在每个 ρ 上把 q 分解成 q=Bf+c,其中 Bf 是最接近的 scalar-compatible component,c 是 cycle-space residual。这样,transitive vs non-transitive 不再是对菌株机制的猜测,而是一个图上 flow 是否落在 incidence matrix image 中的问题。
这个转化的本质区别在于,它改变了信息流组织方式:prior 从几何形状直接跳到 fitness 或 speed,这篇先停在“可观测几何流”,只在图结构上检验 scalar compatibility。新的 inductive bias 是 graph-gradient / cycle-space bias:树状接触图永远不能证伪标量 fitness,只有闭环提供非传递证据。这使方法比单纯 sector-angle fitting 更 generalizable,因为它不要求某个固定生物动力学模型成立;但它也更保守,因为输出本质上只是可见接触上的非势几何残差。
Method
第一,endpoint observability:通过冻结径向前沿假设,把半径当作 deposition order,使单张图像中的边界 trace 可以被视为历史曲线。这解决“单图像没有时间”的问题,但只得到 ρ-clock,不得到 physical time。
第二,boundary-flow extraction:从 log-polar 边界轨迹估计角度导数 q。这个步骤的核心变化是从最终 sector label/count 转向 boundary differential geometry;它关注 pairwise contact dynamics,而不是全局丰度。
第三,graph decomposition:用 incidence matrix B 和权重 W 将 edge flow 分解成 scalar potential part 与 cyclic residual。它解决的是“什么叫无法由标量 fitness 解释”:不是凭视觉直觉,而是看 q 是否在 im(B) 中。cycle residual 的维度 |E|-|V|+1 也直接给出任务上限:没有环就没有证伪能力。
第四,contact design 与统计检验:complete pairwise game 需要足够的 circular boundary walk 覆盖所有边,四型不是六条而是至少八条边界。Gaussian GLR 和 Bonferroni interval scan 提供的是显著性控制,而不是新视觉能力;它们让 residual detection 不至于只是拟合噪声。
Key Insight / Why It Works
最关键的 insight 是把 non-transitivity 表达为 graph cycle-space 中的 residual,而不是表达为某个具体微生物机制。这一点很强,因为它把复杂生态动力学中的不可观测部分剥离掉,只保留 endpoint image 中可检验的代数约束:scalar hierarchy 等价于 edge flow 是梯度场,cyclic dominance 对应闭环和不为零。方法有效的根源不是更强分割器、不是学习表示,也不是 scaling,而是更合适的 latent structure:边界几何 → edge flow → graph Hodge decomposition。
最可能的核心贡献是“可观测量边界 + cyclic residual 检验”这一组合。contact-complete design theorem 也很实质,因为它指出实验设计不是任意多放几个 sector,而是一个闭合 walk 覆盖问题。相反,active design-control、PDE sweep、GPU diagnostics、HMC/Fourier inversion 等部分更像工程延展,和主反问题的逻辑耦合并不强;这些结果可能主要来自 simulation design、compute scaling 和目标函数设定,而不是 radial tomography 带来的新识别能力。
需要直接说:这篇论文的可信部分是 geometry-to-graph inverse problem;较可疑或至少证据不足的部分是从 cyclic residual 继续推到 strain design/control 的链条。文中虽然谨慎写了 governance 和非辨识边界,但 downstream 模块容易给人一种“从一张图到机制再到控制”的错觉。严格看,endpoint 图像只支持 pairwise boundary-flow residual,不能支持机制闭环。
Relation To Prior Work
最接近的谱系是 microbial range expansion geometry、sector boundary stochastic process、graph/Hodge decomposition 和 classical contour-based vision。与 Korolev/Weinstein 一类 sector-geometry 工作相比,差异不是换了图像处理 pipeline,而是把 scalar fitness 从默认解释降级为一个可检验 null model。与 rock-paper-scissors microbial ecology 相比,论文没有提出新的非传递生物机制,而是给出从 endpoint pattern 识别非传递 signature 的反问题形式。
看似新的视觉部分其实多是已有 classical vision 组件重组:固定颜色分割、log-polar remapping、contour tracing、smoothing、GLR test 都不新。实质创新在于三点:一是明确 endpoint 只识别 deposition-coordinate edge flow;二是用 graph-gradient/cycle residual 精确定义 scalar incompatibility;三是把 circular contact design 和 cycle-space 维度联系起来,说明什么图像设计有资格证伪 transitivity。它属于“把领域物理约束转成低维几何统计检验”的路线,而不是 representation learning 路线。
Dataset / Evaluation
evaluation 覆盖了 analytic endpoint、pixel round trip、scalar-null stress、mechanistic frozen-front endpoint、多分辨率合成图和 public-image tracing audit。它对核心 claim 的支持是有限但有针对性的:在合成或受控机制模型下,pipeline 能从像素恢复 flow,并且 cyclic residual 对 programmed cyclicity 有响应;在 scalar-null 下也检查了 nuisance 导致的假阳性。这个设计比只报一个漂亮案例更可信。
但它并没有充分验证真实生物 deployment。真实图像部分更像 provenance/前端 sanity check,不是 raw quantitative microscopy 上的因果验证。最重要的 stress test 反而暴露了边界:在强 center error、bleed-through、anisotropy、extinction 等联合 nuisance 下,标量真值也会被判成 cyclic。这说明 benchmark 支持的是“有 abstention 的受控识别”,不是“单张任意 colony 图像可可靠识别非传递生态”。active-control 评估基本是 simulation-to-simulation,不应被视为支持 endpoint recognition claim 的主要证据。
Limitation
核心前提很重:冻结前沿、径向沉积顺序、中心可估、边界可见且可追踪、sector 不发生拓扑重排、接触图含闭环、重复接触能正确 association。任何一个前提弱化,q 的估计和 c 的解释都会迅速变脆。特别是 center error 和 anisotropic front 可以把纯 scalar flow 投影成 apparent cyclic residual;这不是小噪声问题,而是结构性混淆。
scalability 上限也明显。genotype 数增加后,complete contact design 所需边界数增长,实际 colony 中边界稳定性、sector extinction 和重复边识别会成为瓶颈。图上 cycle-space 维度增加并不等价于信息增加,因为每条边界的几何质量和 covariance 校准会恶化。泛化不是来自数据覆盖或 learned representation,而来自理论约束;这使它在假设内很稳,在假设外会直接 abstain 或误判。
更大的问题是解释边界。论文反复声明不能识别 physical clock、isolated speed 和 mechanism,这是正确的;但后面 active design/control 部分容易把 residual 当成机制 forcing signal 使用。这里增益来源不清,可能主要来自 scaling / simulation objective,而不是 endpoint inversion。文中未充分说明真实实验中如何校准 covariance、如何验证 residual 与实际 pairwise interaction matrix 对齐,以及如何避免 publication-figure preprocessing 带来的 evaluation bias。
Takeaway
- 第一,最值得迁移的是“先定义可观测几何量,再定义不可辨识边界”的姿态;这比直接从图像预测生物参数更可靠。
- 第二,graph-gradient null model 是一个很干净的 inductive bias:凡是 pairwise observable 能组织成图上边流的问题,都可以用 cycle residual 检验是否存在标量潜势解释。
- 第三,实验设计本身是识别能力的一部分。
- 没有闭环接触,任何 endpoint 图像都无法证伪 scalar hierarchy;更多像素或更强模型不能弥补拓扑信息缺失。
一句话总结
这篇论文的实质贡献是把单张径向扩张终态图像中的非传递识别,从模糊的 sector-shape interpretation 重构为一个受可辨识边界约束的 graph edge-flow decomposition 问题;它是领域物理约束驱动的几何统计方法,而不是视觉模型或生物机制发现方法。