精读笔记
Problem Setting
论文处理的是从常规 H&E 切片直接生成空间基因表达的问题,但真正问题不是“图像到向量”的监督学习,而是弱形态信号下的结构化条件分布恢复。H&E 中可见的是细胞形态、组织结构和局部微环境,表达谱中包含大量不可见或弱可见的状态变量;因此同一类形态可能对应多个表达模式,点回归天然会向条件均值收缩。此前方法卡在两个方向:回归方法推理快但过平滑,生成方法能描述分布但推理慢;同时输出基因被当成独立维度处理,忽略 co-expression、pathway coupling 和不同基因的信息密度差异。这个任务的关键矛盾是:如何在没有额外 wet-lab 信息的情况下,用单步推理得到不像回归均值那样塌缩、又不需要多步采样的表达预测。
Motivation
已有路线不够的地方在于它们主要优化 per-gene error,而不是显式塑造预测表达分布。对于 ST 预测,低 MSE 不等于保留空间异质性;很多下游任务关心的是 cluster boundary、marker localization、rare population 和空间结构。作者的核心观察是:输出端本身有强结构,真实表达分布可以反过来指导模型如何生成,而不是只作为逐点标签。关键缺口是一个训练时有生成分布约束、推理时仍保持一次 forward 的机制;以及一个能把 gene-gene dependency 注入输出过程的架构。DriftST 的方向可以理解为:用 training-time distribution transport 取代 test-time iterative generation,用 gene-aware output modeling 取代普通多头回归。
Core Idea
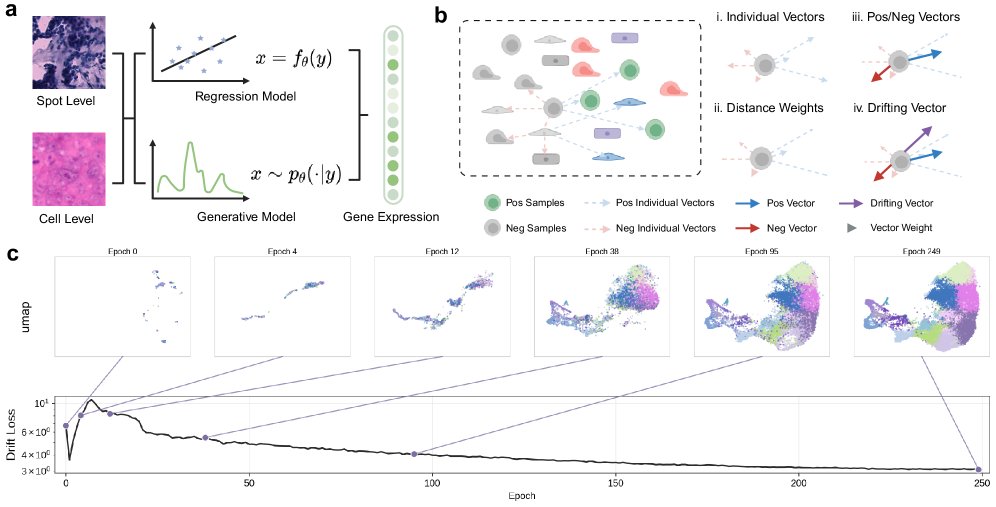
DriftST 的核心思想是把 H&E-to-ST 从回归问题改写成条件 pushforward distribution matching:图像 encoder 和生成器定义一个从 H&E patch 到表达空间的映射,训练时通过 cellular drifting field 推动预测分布靠近真实 ST 分布。这个 drift field 由 paired ground truth 的吸引和其他预测样本的排斥组成,避免所有样本向全局均值坍缩;multi-scale kernel 则让局部表达邻域和全局分布形状都参与训练。
它和 diffusion / autoregressive 生成模型的本质区别是:不在推理时沿路径逐步采样,而是在训练时把分布演化压进参数里。推理时模型已经学成一个“直接落到表达流形附近”的映射,所以是 one-step。新增的 inductive bias 来自两端:目标函数端用表达分布几何约束预测,架构端用 co-expression prior 和 gene-wise gating 重新组织输出信息流。更 scalable 的地方主要在 inference;training 并不轻,因为需要 dropout copies、prediction bank 和额外 drift computation。
Method
第一,Cellular Drifting 解决的是回归均值塌缩。paired expression 提供吸引方向,历史预测和同 patch stochastic predictions 提供排斥方向;分开归一化 positive 和 negative 权重,避免大量 negatives 淹没唯一 positive。核心变化是目标从“贴近一个标签点”变成“在表达空间中移动到更合理的局部分布位置”。
第二,prediction bank 解决 mini-batch 负样本覆盖不足。它复用历史预测来近似当前模型生成分布,使 drift field 不只依赖当前 batch 的偶然结构。这更像 memory-based distribution regularization,而不是经典生成采样。
第三,STransformer 解决输出维度无结构的问题。gene embedding + FiLM 把共享形态特征拆成 gene-specific tokens;co-expression attention 用训练集相关矩阵作为 attention bias,使基因间依赖进入预测过程;gene residual gate 让不同基因接受不同程度的 refinement,避免把 housekeeping-like genes 和高度可变 marker genes 用同一计算路径处理。
第四,ZINB 辅助项解决 count data 的分布约束问题。它不是论文最核心的创新,更像防止纯 drift 几何目标偏离计数生成机制的正则化。
Key Insight / Why It Works
最可能真正有效的是 drift loss + prediction bank 形成的分布级正则,而不是单个网络模块。H&E-to-ST 的监督信号噪声很大,单点 MSE 会奖励平均化;drift field 通过“靠近 paired target、远离已有预测模式”给模型一个 anti-collapse signal,迫使预测在表达空间保持一定分散性。这解释了为什么它在 PCC、JSD、SSIM 和 downstream clustering 上比纯回归更好。
第二个有效来源是 output-side inductive bias。基因表达不是独立标签,多数 baseline 把 gene dimension 当成普通输出通道,等于让模型从数据中重新发现 co-expression structure。DriftST 直接把训练集 co-expression matrix 注入 attention,尤其对低变异、长尾或弱形态可见基因有帮助。但这也意味着模型可能在利用 dataset-level gene correlation 补全不可见信息,而不是真正从 H&E 读出了这些基因。
第三,部分增益可能主要来自 scaling / representation alignment。CONCH + UNI2 的 frozen pathology features 很强,邻域上下文也提供空间平滑和 tissue niche 信息。Table 4 显示换成 ResNet 后明显下降,说明视觉 foundation encoder 是性能底座。文中未充分说明在同等 encoder、同等邻域、同等训练预算下,drift objective 相比强 ZINB / contrastive / distributional regression baseline 的净收益。
第四,gate 的贡献可能被高估。ablation 中去掉 gate 后指标下降存在但不大,且附录里 gate 超参变化几乎不影响性能。这说明 gate 更像稳定训练和轻量 gene-wise capacity allocation,而不是决定性机制。真正核心更可能是 drift + co-expression prior + strong encoder。
需要警惕 implicit memorization。prediction bank 存历史预测,本身不是存 ground truth,但在空间 CV 或单 slide 场景下,邻近 patches 与表达模式高度相关,模型可能通过形态/空间邻域学习到 dataset-specific expression atlas。论文没有充分排除 benchmark overlap 或 spatial autocorrelation 带来的高估。
Relation To Prior Work
它最接近三条路线的交叉:H&E-to-ST regression、conditional generative modeling、drifting / flow-like distribution transport。相对 ST-Net、HisToGene、Hist2ST、BLEEP、M2ORT、TRIPLEX 这类方法,DriftST 的本质差异不是用了更复杂 backbone,而是训练目标从 pointwise regression / alignment 转成 prediction distribution shaping。相对 STEM diffusion 和 GenAR autoregressive,它保留生成式分布匹配的诉求,但取消 test-time iterative decoding,把生成成本搬到训练期。
看似新的部分中,co-expression attention 本质上是 graph-biased / prior-biased attention 在基因维度的应用,gene residual gate 也是 conditional computation / residual gating 的迁移;这些是合理重组,不是概念上全新。实质创新在于把 drifting model 改造成 paired conditional prediction setting,并用 positive attraction + negative prediction bank 来避免条件均值塌缩。这篇属于“训练时分布对齐、推理时直接映射”的技术谱系,介于 regression 和 diffusion 之间。
Dataset / Evaluation
评估覆盖了 spot-level 和 cell-level、不同平台和组织,足以初步支持 unified framework 的 claim。尤其同时在 Xenium cell-level 和 Visium / SpatialTranscriptomics spot-level 上比较,说明方法不是只为单一分辨率调参。downstream 分析也比只报 PCC 更有意义,因为 cell-type recovery、marker spatial map、JSD/SSIM 更接近真实生物使用场景。
但 evaluation 仍有明显限制。cell-level 是单 slide 的 spatial 5-fold,本质上测试的是 slide 内空间外推,不等于跨患者泛化;spot-level leave-one-slide-out 更接近真实 domain shift,但 Kidney Visium 已暴露出 batch / patient heterogeneity 下性能下降。benchmark 没有充分验证跨机构、跨染色、跨扫描仪、跨 disease state 的 robustness。另一个问题是 baseline 是否都用了同等级 pathology foundation encoders、同样邻域信息和同样训练预算,文中未充分说明;如果没有,SOTA claim 中会混入 representation scaling 的收益。
Limitation
DriftST 的上限首先来自可见性:H&E 并不包含完整 transcriptome 信息,很多表达差异在形态上不可辨。模型可以利用 co-expression prior 和数据集统计补全,但这更像 conditional imputation,不是从图像直接观测到表达。对于 rare immune states、activation states 或 disease reprogramming,形态-表达映射弱时仍会失败。
第二,方法依赖训练 panel。它只能预测训练集中定义的 200–300 个 HMHVG / panel genes,不能自然扩展到全转录组;co-expression prior 也是 panel 内估计的,换 panel 就要重建输出空间。所谓 unified 更多是 spot/cell resolution 上的统一,不是 gene space 或 platform biology 上的统一。
第三,泛化仍不充分。Kidney Visium 的结果已经说明跨 patient、batch effect、临床状态差异会削弱模型。co-expression matrix 在正常组织和肿瘤异常区域可能不同,作者用 spot-dependent scaling 调节 prior,但文中未充分证明它能处理真实 regulatory rewiring。
第四,one-step generative inference 有一定表述风险。推理确实单步,但训练依赖 K 次 dropout、prediction bank 和较长训练;这不是免费生成,而是把多步采样成本转移到训练。若目标场景需要频繁重训或跨域适配,成本优势会变小。
第五,增益归因不清。dual foundation models、空间邻域、ZINB、drift、co-expression、gate 同时存在;ablation 虽有帮助,但没有完全隔离强 encoder scaling 和 generative objective 的贡献。部分提升可能主要来自 scaling / data coverage。
Takeaway
- 第一,H&E-to-ST 未来不应只看 per-gene regression error,核心是如何保持表达分布、空间结构和下游 cell-state 可用性。
- 第二,单步生成的可行路径不是一定要 distill diffusion,而可以在训练时做 distribution transport / anti-collapse regularization,让推理仍保持直接映射。
- 第三,输出空间结构很重要。
- 对于高维生物预测任务,把基因、蛋白、peak 或 pathway 当作 structured tokens,显式注入 co-expression / regulatory prior,可能比继续堆图像 encoder 更有迁移价值。
一句话总结
DriftST 是 H&E-to-ST 方向中把生成式分布建模训练化、把推理回归化的一步,真正贡献在于用 drifting-style distribution regularization 和 gene-structured output bias 缓解回归均值塌缩,而不是单纯提出一个更大的预测网络。