精读笔记
Problem Setting
论文标题:Micrometer-sized molecular robot changes its shape in response to signal molecules(Science Robotics / 2017)。
这篇论文真正处理的是“分子机器人系统集成”的最小闭环雏形:不是单独证明 DNA 可以计算、kinesin 可以驱动、liposome 可以变形,而是证明这三者能在同一个微米级封装体内形成一条从 molecular signal 到 mechanical morphology 的因果链。
真正困难点不在某个模块,而在跨尺度接口:DNA 信号反应是局域、扩散受限、纳米尺度的;kinesin-microtubule 产生的是沿 filament 的相对滑动;liposome 形变需要克服膜张力、弯曲能和流体膜耗散。以前方法常卡在两端之一:要么是 DNA nanorobot 级别的结构/逻辑,缺少可见的微米尺度力学输出;要么是 active vesicle/人工细胞模型,能形变但不可编程、不可由特异分子信号控制。
关键矛盾是:分子器件越可编程,通常越弱、越慢、越受环境影响;执行器越有力,通常越难以精确接入逻辑控制。作者选择的解法不是增强 DNA 的机械输出,而是让 DNA 只负责控制“力是否能传到膜”。这是这篇文章的核心问题定义。
Motivation
已有路线不够的地方在于缺少一个通用的 molecular control-to-actuation interface。单分子机器人受 Brownian motion 支配,行为随机且难以形成协同;liposome active matter 可以产生细胞样形变,但更像自组织材料,缺少外部或内部 signal 对执行器状态的选择性控制;DNA computing 虽然可编程,但单独存在时难以产生微米尺度形态响应。
作者的关键观察是:细胞并不是让每个分子器件单独完成“机器人行为”,而是把 sensing、processing、actuation 放进同一个 compartment,并通过耦合关系改变整体状态。对应到工程实现,最缺的不是更复杂的 DNA circuit,而是一个可以把 DNA 序列识别转成 mechanical coupling 的 clutch。
因此这篇论文的动机是补上“programmability 到 motility/shape change”之间的接口缺口。它不追求复杂控制策略,而是先证明一个最小系统:特定信号分子可以决定执行器是否对 body 做功。
Core Idea
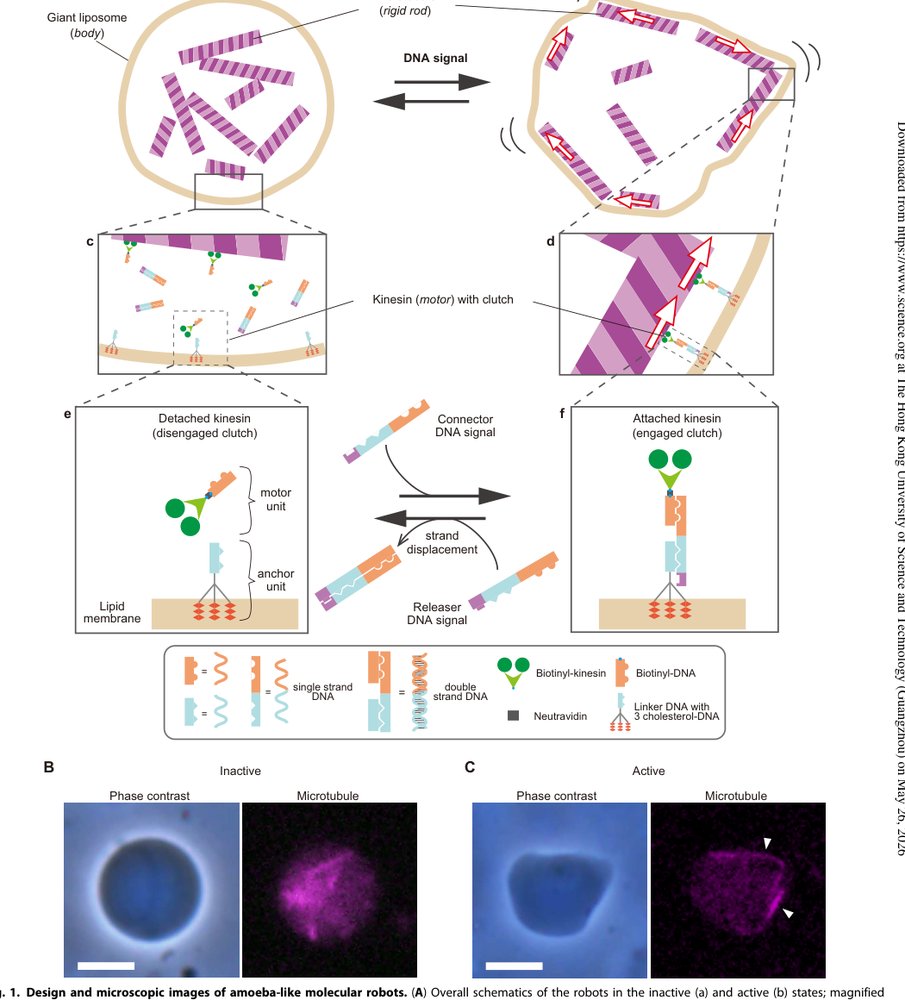
核心思想是把分子机器人建模成 body-actuator-controller 的耦合系统,其中 controller 不直接驱动形变,而是控制 actuator 和 body 之间的力传递通道。这个建模变化很关键:DNA 的优势是序列可编程和反应可设计,不是高功率机械输出;kinesin/microtubule 的优势是持续消耗 ATP 产生运动,不是可编程逻辑。把两者串起来的最自然接口就是 clutch。
具体说,DNA strand displacement 被用作“机械离合器”的状态切换机制:connector DNA 让 kinesin motor unit 与 cholesterol-DNA anchor unit 连接,anchor 插入膜后,microtubule-kinesin 的滑动对膜施力;releaser DNA 则通过链置换断开连接,使马达仍可能活动但不再对膜产生有效形变。这使得 molecular signal 控制的是 coupling topology,而不是控制动力源本身。
与 prior 的本质区别在这里:它不是又一个 DNA device,也不是又一个 active vesicle,而是把 DNA reaction network 放到了 actuator-body interface 上。这引入的 inductive bias 是“可编程性应作用于耦合关系”,而不是作用于材料本体或执行器能量供应。这一点比论文里的具体 DNA 序列或膜配方更值得迁移。
Method
1. Giant liposome 作为 body:它解决的是单分子尺度行为过度随机、无法形成协同输出的问题。微米级 compartment 允许多个 motor、microtubule、DNA device 在同一边界条件下工作,使局部随机事件在整体形态上被平均成可观测状态。但这个 body 不是被动容器,它的膜流动性、相分离和力学模量直接决定执行器输出是否能显现。
2. Kinesin-microtubule 作为 actuator:它解决的是 DNA 本身机械功率不足的问题。kinesin 消耗 ATP 沿 microtubule 产生滑动,若被锚定到膜内侧,就能把相对运动转成膜形变。这里需要注意,执行器的存在并不自动导致机器人行为;只有力传递路径建立后,运动才被投影到 body morphology 上。
3. DNA clutch 作为 controller-actuator interface:它解决的是“执行器可控性”问题。motor unit 和 anchor unit 之间的连接由 connector/releaser DNA 控制,strand displacement 提供序列特异的开关逻辑。这个设计的核心变化是把 DNA 从信息分子变成机械耦合状态的调度器。
4. Photoresponsive DNA signal:它解决实验上在封装体内部定时释放信号的问题。UV 裂解 hairpin prDNA 产生 connector 或 releaser,使作者能在观察过程中触发状态变化。这个机制主要是实验控制手段,不是机器人最终形态下最本质的 sensing 机制;真实环境感知仍未实现。
5. 膜组成优化:DOPC/DPPC/cholesterol/PEG-lipid 的选择本质上是在调节膜的流动性、相分离和非特异吸附。DPPC 可能形成相对 rigid domain,帮助 cholesterol-DNA anchor 传递力。这里更像关键 engineering enabling condition,而不是新的控制原理。
Key Insight / Why It Works
最重要的 insight 是:在分子机器人里,控制执行器的有效方式可能不是调控 motor activity,而是调控 motor 与 body 的 mechanical coupling。kinesin 一直可以消耗 ATP,microtubule 也一直存在;active/inactive 的差别不是动力源有没有,而是动力源是否被接到膜上。这一点解释了为什么一个简单的 DNA strand displacement 就能放大成微米尺度形态变化:DNA 反应只需改变连接状态,后续能量由 ATP-driven motor 持续提供。
方法有效的第二个原因是 compartmentalization。所有组分被封装在 liposome 内,局部浓度、边界、膜锚定位点和执行器共同形成一个反应-力学耦合场。单个分子事件不需要精确定位,只要足够多 clutch 处于 engaged 状态,microtubule gliding 就会在膜上产生持续扰动,表现为整体形态波动。
第三个原因是膜不是完全流体化的惰性边界。文中显示纯 DOPC/cholesterol/PEG-lipid 条件下形变效率差,加入 DPPC 后连续形变 yield 改善。作者推测相分离/刚性 domain 提供 foothold。这一点很可能是实验能否成立的关键,但机制没有被充分定量。换言之,论文的系统成功不只是 DNA clutch 的成功,也依赖一个合适的 soft-matter operating regime。
最可能的核心贡献:DNA-controlled clutch 作为 signal-to-mechanics transducer。最可能只是辅助但不可缺的部分:photoresponsive trigger、膜配方筛选、抗氧化/ATP regeneration 等体系工程。这里不存在机器学习意义上的 scaling/data/retrieval;如果用抽象术语类比,它更接近 better inductive bias 和 representation alignment:把 DNA sequence state 对齐到 mechanical coupling state,把 motor activity 对齐到 membrane deformation。
需要直接指出的是,文章中“robot”一词偏强。当前系统没有环境 sensing,没有内部决策,没有可逆多步控制,也没有目标导向 locomotion。它证明的是可编程分子信号可以门控 active material 的形态状态,而不是实现了完整机器人行为。
Relation To Prior Work
最接近的谱系有三条:DNA nanotechnology / DNA computing,kinesin-microtubule active matter,以及 artificial cell / giant liposome reconstitution。
相对 DNA nanorobot 工作,这篇的不同点不是 DNA 结构更复杂,而是 DNA 不再主要承担结构折叠、cargo release 或逻辑计算,而是作为 clutch 控制力学耦合。它把 DNA 的序列可编程性外包到一个机械接口上,这是实质新增信息。
相对 Keber、Loiseau 等 active cytoskeletal vesicle,这篇的不同点是可控性。prior 已经说明 microtubule/kinesin 能在 vesicle 内诱导 protrusion、blebbing 或 active deformation,但那些行为更多是自组织动力学。本文新增的是特异 DNA signal 能决定 active system 是否作用到 membrane。
相对 Wollman et al. 这类 DNA-programmed motor/protein assemblies,本文的推进在于把 motor-DNA 控制放进微米级 compartment,并把输出读成 body morphology,而不是仅在开放溶液或表面体系中观察 assembly/disassembly。
看似新的部分里,liposome 制备、strand displacement、cholesterol-DNA membrane anchoring、kinesin gliding assay 都是已有工具重组;实质创新在系统层级的接口设计:molecular signal → clutch state → force transmission → membrane shape。
Dataset / Evaluation
这里没有 dataset,评价是湿实验 proof-of-concept。任务覆盖范围非常窄:单一 body 类型、单一 actuator 类型、两种 signal-triggered switching 方向、单个或少量 robot 的形态时间序列。它是真实物理系统而非仿真,因而对“能否实际集成”这个 claim 有较强支持。
核心评价指标是形态半径随角度和时间的变化,以及 radius SD 的 active/inactive 区分。这个评价足以证明 engaged/disengaged clutch 对形态动力学有影响,但不足以证明更强 claim,例如 programmable motility、general molecular robotics platform 或可扩展控制架构。
实验设计中的强点是对 active/inactive、ATP 缺失、signal premixing、UV-triggered prDNA 和 gel electrophoresis 都做了基本验证,因果链相对完整。弱点是统计和系统异质性展示有限,机器人 yield 约 10% 且个体差异没有被系统建模;switching 时间远慢于裸 DNA 反应,说明在真实封装环境下 controller 与 actuator 的耦合效率仍是瓶颈。
因此 evaluation 支持“分子信号可门控形变行为”这个中心结论,但不支持“已经具备复杂机器人功能”。
Limitation
1. 不可逆控制是硬限制。当前 connector/releaser 通过 strand displacement 和 photocleavage 实现一次性切换,不能反复循环。没有 reversible clutch,就很难谈真正的控制策略或时序程序。
2. 形变无方向性。active state 只是连续非球形波动或旋转式形态变化,不能产生稳定净位移。所谓 amoeba-like 更像形态表象,而不是具备 amoeboid locomotion 的功能等价物。
3. 可扩展性依赖膜-蛋白-DNA 的复杂环境。作者承认 DNA strand displacement 在 liposome 中本可约 300 s 完成,但机器人 switching 约 700 s 后才完成。这说明瓶颈不在 DNA 反应本身,而在 tethered complex diffusion、膜固定反应、静电相互作用和机械松弛。复杂 DNA circuits 接入后,速度和泄漏问题很可能更严重。
4. 力学机制未被充分定量。DPPC domain 被认为提供 foothold,但文中没有直接测量 anchor 分布、domain localization、局部膜刚度与形变效率之间的因果关系。增益来源不清,可能相当一部分来自膜配方筛选和体系工程。
5. 平台性 claim 仍偏乐观。论文说可冷冻邮寄、可让其他实验室安装自定义分子器件,但这不等于系统可模块化扩展。每个新分子器件都会改变离子条件、拥挤度、膜相互作用和非特异吸附;“plug-and-play”在文中未充分说明。
6. 控制问题被转移了而非解决了。本文把复杂控制简化为 clutch 开关,这是合理的第一步,但后续若要 chemotaxis,需要外部 sensing、跨膜 signal transduction、内部 computation、空间极性建立和 actuation asymmetry。当前系统只解决其中最靠后的 actuator gating。
Takeaway
- 1. 这篇真正推动的是 molecular robotics 的接口思想:把 DNA 的可编程性放在 force-transmission layer,而不是让 DNA 本身承担机械输出。
- 2. 对人工细胞/active matter 方向,关键启发是 active cytoskeleton 需要可编程 clutch 才能从自组织材料走向可控机器人;控制 coupling topology 可能比控制每个 motor 更可行。
- 3. 对后续工作,最值得做的不是堆更多 DNA 模块,而是解决三件事:可逆 clutch、空间非对称性、跨膜 sensing 到内部 actuation 的闭环。
- 4. 这篇的成功很大程度来自正确的跨尺度分工:DNA 做识别和状态切换,protein motor 做能量转换,lipid membrane 做可变形 body。
一句话总结
这篇论文在 molecular robotics 中的定位是一个最小系统级 proof-of-principle:它用 DNA strand displacement 构造机械离合器,把序列特异信号转译为 kinesin-microtubule 对 liposome 膜的力传递开关,从而首次展示了可编程分子信号门控的微米级形态变化。