精读笔记
Problem Setting
论文标题:Toward a living soft microrobot through optogenetic locomotion control of Caenorhabditis elegans(Science Robotics / 2021)。
这篇论文真正处理的问题是:如何把一个完整活体线虫从“自主动物”转化为“可工程控制的柔体微机器人”。关键不是证明光遗传能让肌肉收缩,这在 C. elegans 里已有基础;关键是能否在切断内源神经控制之后,用外部控制重建足够正确的分布式肌肉激活模式,使整个身体产生稳定推进和可控转向。
真正困难点在于 C. elegans crawling 是一个连续体力学问题:推进来自身体曲率波、肌肉激活波、基底摩擦/支撑之间的相位组织,而不是来自某个局部 actuator 的力输出。传统微型机器人卡在微尺度高效 actuator、能量密度、柔体结构制造和控制一体化;已有 biohybrid microrobot 多利用细菌/精子/组织片段的单一推进能力,控制通常发生在方向层面,而不是重新编排一个多细胞动物的 body-level locomotion。
因此这篇的关键矛盾是:活体动物本身有复杂、强耦合、难建模的运动生成系统;但机器人控制需要可观测、可寻址、可闭环调节的输入输出结构。作者的做法是直接绕开神经系统,把动物身体保留下来,把控制权转移到外部光学闭环。
Motivation
已有两条路线都不够。仿生微机器人路线试图复刻生物结构和运动,但在微尺度上 actuator、能源和制造复杂度很快成为瓶颈,最后往往只能得到高度简化的形态,性能和真实生物差距明显。biohybrid 路线虽然借用了活体推进,但多数系统只是把生物体当成 propulsion module,缺少对多自由度身体协调的可编程控制。
作者的核心观察是:C. elegans 已经拥有适合微尺度运动的柔性身体、肌肉执行器和能量供应;真正需要替换的是“谁来产生肌肉时空激活模式”。如果能关闭内源神经输入,同时让肌肉保持光敏可激发,那么可以把线虫身体视为一个自然制造的 distributed soft robot,并由外部视觉-控制-光刺激回路接管运动。
关键缺口在于:过去 optogenetics 更多用于扰动神经/肌肉以研究行为机制,而不是把整套 locomotion control loop 工程化;过去微机器人控制也很少能在完整多细胞动物身体上实现可编程 gait generation 和导航。
Core Idea
核心思想是把 C. elegans 的运动控制从“神经系统生成行为”改写成“外部系统在活体连续体上写入肌肉激活波”。这里的建模变化很重要:线虫不再主要被看作一个待解释的生物神经系统,而是被看作一个已有形态和材料优势的柔性机器人平台;肌肉是 distributed actuator,身体曲率是状态,patterned light 是可编程输入,显微视觉是外部传感器。
论文引入的关键 inductive bias 是相位差:不是直接追踪自然神经活动的全部细节,也不是拟合复杂神经-肌肉模型,而是抓住“肌肉活动波相对身体曲率波错位”这一低维力学变量。这个 bias 使控制问题从高维、个体差异极大的生物系统,压缩成可以用扫描光图案实现的时空相位控制。
和 prior 的本质区别是控制层级不同。细菌/精子 microrobot 多是在已有推进器上做 steering;这里是对一个完整柔体动物的 body wave generation 进行外部重构。它不是简单“用光驱动活体”,而是把 locomotion pattern 的生成机制抽象成相位关系,再把该相位关系写回瘫痪身体。
Method
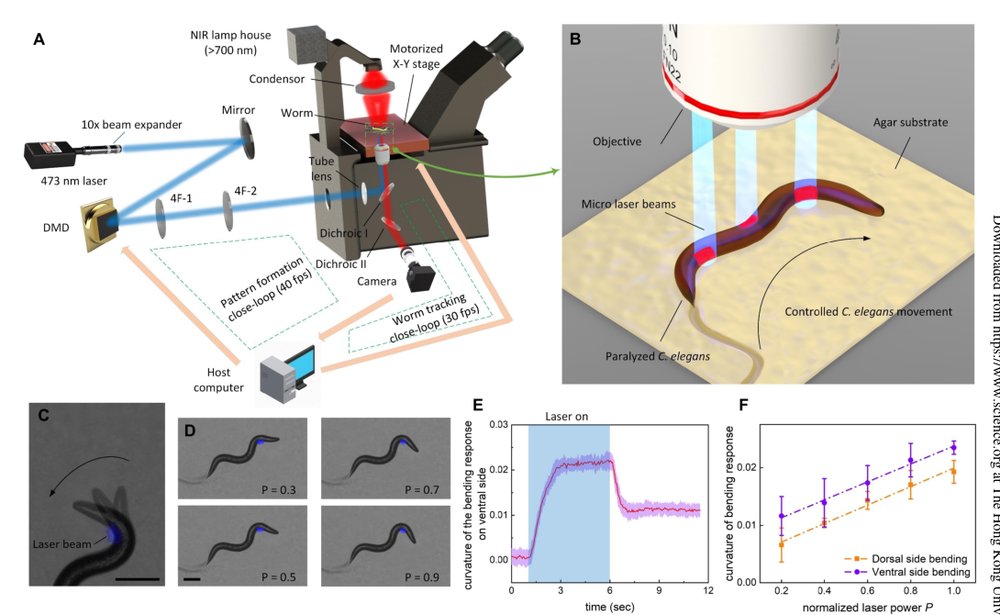
1. 切断内源 sensorimotor loop:ivermectin 使神经系统超极化,让线虫停止自主爬行,同时肌肉仍可被 ChR2 光激活。这个步骤的作用是消除内源运动命令与外部命令的冲突,把控制权交给工程系统。代价是它并不干净,也会部分影响肌肉,这是后续成功率低的根源之一。
2. 将肌肉变成空间可寻址 actuator:myo-3 promoter 下肌肉表达 ChR2,DMD 生成 473 nm patterned light,选择性照射 dorsal/ventral 身体段。机制层面的意义是把连续身体离散成一串可编程肌肉段,而不是依赖自然 motor neuron pattern。
3. 用相位差设计 gait:自然 crawling 中肌肉活动与身体曲率存在相位错位;作者用扫描的二值正弦条纹在瘫痪身体上重建这一错位。这里的核心不是二值化本身,而是让激活波从 head 到 tail 扫描并与身体变形形成稳定相位差,从而积累推进力。
4. 用头部不对称光强控制方向:身体后段主要提供推进,头部半波长区域决定后续轨迹方向。调节 dorsal/ventral 激活强度可以改变 head bending,从而实现 shallow turn、gradual turn、omega turn 等 motif。这里相当于把转向控制降维为 head-region gain modulation。
5. 用外部视觉闭环做导航:图像处理实时估计中心线、曲率、质心和运动方向;底层维持图案投影与身体对齐,顶层用预测 P 控制调节下一周期光强,使运动方向朝目标点收敛。动态模型没有直接用于控制器设计,控制器本质是经验几何模型 + 视觉反馈纠偏。
Key Insight / Why It Works
最重要的 insight 是:serpentine crawling 的可控性主要由肌肉激活波与身体曲率波之间的相位关系决定。论文通过能量推导给出直观解释:当肌肉 torque 与曲率完全对齐或反相时,主要是在维持/抵消形状,不产生净推进;只有存在适当相位差,肌肉做功才能转化为沿身体前进方向的 thrust。这个结论把复杂的 neuromechanical control 压缩成一个可外部写入的 phase-control 问题。
方法有效的真正原因不是 DMD 分辨率高,也不是 P controller 多聪明,而是作者找到了一个低维、物理上充分的控制变量。扫描光图案本质上是在模拟自然 motor program 的 traveling activation wave;只要肌肉还能响应、摩擦环境足够稳定、身体形态能被实时跟踪,线虫身体自身的柔顺性和基底接触会完成剩余的力学计算。这是典型的 morphological computation + externalized control。
最可能的核心贡献有两个:第一,实验证实相位差可以作为 crawling thrust 的工程控制变量;第二,把完整 C. elegans 的 locomotion loop 外部化,实现了从 paralysis 到 controllable gait 的重构。相比之下,40 fps、多线程、DMD intensity compensation 等主要是必要 engineering,不是概念贡献;闭环导航展示也更多是系统集成能力,不是新的机器人规划算法。
这不是 scaling、不是 data coverage、不是 learning,也不是 retrieval。它更接近 better inductive bias:用 biomechanics 找到正确的低维 latent structure,再用 optogenetics 写入该结构。控制器没有形成长期状态建模,所谓通过迷宫主要是外部视觉系统给出 waypoint 和即时方向误差后的反馈调节,不应被理解为 RoboWorm 自身具备空间认知。
Relation To Prior Work
与 C. elegans optogenetics 系统如 CoLBeRT、projector-based illumination 最接近的部分是实时追踪和模式光刺激;但那些平台主要用于神经科学扰动,目标是解释行为回路。本文把同类工具转为机器人控制接口,核心新增是将 optogenetic stimulation 组织成闭环 gait generation。
与 biohybrid microrobot 相比,本文不是借用单个细胞或组织作为推进器,而是保留一个完整多细胞动物的柔体结构和肌肉分布。细菌磁控、精子微锥、Tetrahymena 等系统通常控制方向或群体行为,推进 pattern 由生物体自身生成;RoboWorm 则切断内源推进生成,并外部重写肌肉时空模式。这是本质差异。
与仿生柔体微机器人相比,本文的创新不是制造出新的人工材料,而是利用活体身体绕过制造难题。其技术谱系可以看作:optogenetic neuromodulation → biohybrid actuation → externalized closed-loop animal locomotion control。看似新的“living soft microrobot”概念,实质上是已有 optogenetic manipulation、worm biomechanics、视觉伺服控制的重组;真正实质创新在于用相位差理论把这些组件连成一个可工作的 locomotion controller。
Dataset / Evaluation
评估覆盖了三层 claim:肌肉段响应、gait 生成、闭环导航。肌肉响应实验说明光强可调且 dorsal/ventral 响应不对称;自然 crawling calcium imaging 和动力学分析支持相位差假设;瘫痪线虫上的前进、转向、omega turn、目标点导航和迷宫通过支持“外部控制可重建 locomotion”这一主张。
但 evaluation 的外推范围有限。实验场景基本是 agar plate 上的单虫、显微镜视野内、外部投影和外部视觉辅助;没有展示复杂三维微环境、液体环境、真实 cargo manipulation、长时间部署或多虫控制。迷宫实验更像 waypoint-following demo,验证的是闭环系统可以纠偏,而不是线虫具备自主导航。
核心 claim“能把瘫痪 C. elegans 转成可控 living soft microrobot”被支持;更强的应用 claim,如用于 micromanipulation、复杂微环境导航、体内部署,目前证据不足。尤其成功率只有约 7% 这一点意味着系统可重复性和可扩展性仍是主要问题。
Limitation
1. 生物执行器一致性是硬上限。ivermectin 并不是只关掉神经,它也影响部分肌肉;不同个体、不同身体段、dorsal/ventral 之间响应差异显著。论文自己报告成功率只有 5–10% 量级,这说明控制方法依赖一小部分肌肉响应足够均匀的个体。
2. 控制窗口很短。单个 RoboWorm 有效实验约 10–15 分钟,可能受肌肉疲劳、离子浓度平衡、ATR 消耗等限制。文中未充分区分这些因素,因此长期部署能力不清楚。
3. “untethered”容易误读。线虫身体没有物理 tether,但控制完全依赖显微镜、DMD、外部视觉和电动平台;从系统角度看,这是高度 tethered 的外部控制环境。它不是自治微机器人。
4. 泛化依赖环境稳定性。相位差推进模型建立在 agar crawling、可提供稳定支撑/摩擦的平面基底上。换成非均匀摩擦、液体、复杂障碍接触或体内软组织环境后,推进机制和控制律未必保持。
5. 闭环控制较浅。顶层控制是几何预测 P control,不包含长期状态、接触规划或不确定性建模。迷宫通过主要来自预设中间目标和视觉纠偏,不应被解释为 planning 能力。
6. 后退控制失败暴露了身体可寻址性的边界。论文提出反向扫描理论上可实现 backward crawl,但尾部无法像头部那样有效响应,实际没有完成。这说明“相位差足够”仍受生物执行器空间分布和局部肌肉能力限制。
Takeaway
- 1. 这篇真正值得记住的是:对于连续体生物 locomotion,找到正确的低维力学相位变量,比试图复刻完整神经控制更重要。
- 相位差是一个可迁移的控制抽象。
- 2. 活体微机器人路线的优势不在“更智能”,而在把复杂制造、柔体材料、能量供给和微尺度 actuator 问题外包给生物体;工程系统只需要接管信息流和控制变量。
- 3. 未来真正值得做的是更干净的神经 silencing / 肌肉保真激活、更高成功率的个体控制、在非理想环境下的接触-摩擦自适应,以及把外部控制逐步内嵌或半自主化。
一句话总结
这篇论文在 biohybrid microrobotics 中的真正贡献,是把 C. elegans 从自主动物重构为外部视觉-光遗传闭环控制的分布式柔体执行器,并证明“肌肉激活—身体曲率相位差”可以作为活体线虫爬行控制的核心力学接口。