精读笔记
Problem Setting
论文标题:Modular reconfiguration of DNA origami assemblies using tile displacement(Science Robotics / 2023)。
这篇论文的真实问题不是“做一个 sword 变 snake 的 DNA origami 演示”,而是:DNA origami tile assembly 是否能拥有类似 strand displacement 的可编程替换原语,从而支持模块级结构重配置。已有 tile assembly 可以编码形状,已有 dynamic DNA devices 可以做局部构象变化,但在 tile assembly 尺度上,要让一个已装配模块被另一个复杂模块替换,并让多个替换在同一体系中并行发生,仍然缺机制。
困难点在于 tile 不是短链 DNA。tile edge 的相互作用强、空间尺度大、柔性有限,branch migration 需要两块大结构同时弯曲/呼吸;同时 active edge 之间很容易出现 spurious binding 或 self-occlusion。关键矛盾是:为了稳定装配,需要较强 tile-tile interaction;为了可重配置,又需要足够弱和可逆的局部接触来允许 branch migration。以前的 tile displacement 只在小阵列、简单 invader、相同初末形状上证明过,尚不足以说明它能成为 modular reconfiguration 的通用机制。
Motivation
已有路线缺的是“结构层级的动态信息流”。DNA origami 过去更多是把 addressability 用于静态定位、形状编码、局部机械运动,或者用 assembly/disassembly 控制状态;这些方法要么缺少隐藏 binding domain 和级联能力,要么 specificity 依赖温度窗口,难以在室温和浓度扰动下保持正交。
作者的核心观察是:strand displacement 的威力并不只在 DNA 序列,而在反应组织方式——toehold 触发、branch migration、sequestered domain activation、动力学可调。如果能把这个抽象原语提升到 tile 边缘尺度,就可能把 DNA origami assembly 从静态图案编程变成可重配置的模块系统。关键缺口是:tile displacement 是否能支持足够大的 reaction design space、复杂 invader 的可制造性,以及多个反应的低串扰并行。
Core Idea
论文真正的核心思想是把 DNA origami tile 的边缘从“静态粘接接口”重定义为“可被 invasion 激活的反应域”。invader 不是简单与目标 tile 结合,而是通过 toehold 进入一个竞争态,使原 cover assembly 和 invader assembly 在同一 base edge 上进行 branch migration。这样,结构变化被表示为 reaction pathway,而不是热平衡下的 assembly preference。
与 prior 的本质区别在于信息流从全局 annealing / melting 转移到局部 trigger-controlled replacement:旧模块的释放不是通过整体解离,而是被新模块沿边缘逐步替换。这引入了一个重要 inductive bias:重配置只能从暴露 toehold 开始,且只能沿设计好的 migration domain 传播。因此系统的 specificity 来自路径约束,而不只是 pairwise binding energy。理论上这更 scalable,因为可以把复杂结构变化拆成局部替换反应;也更 generalizable,因为 invader 可以是单 tile、多 tile、变长 polymer 或 3D barrel 的 opening trigger。
Method
1. mixed giving/receiving domain:解决的是反应 identity 不够用的问题。原先 giving-to-giving 类型把 toehold 和 branch migration 的 edge identity 绑得太死,可设计反应数很小。混合 G-R / R-G 让 toehold identity 与 migration identity 部分解耦,核心变化是把同一个 branch migration domain 与多个 toehold 组合,扩大正交反应空间。
2. discontinuous toehold:解决的是 toehold 位置组合受限的问题。作者利用 origami edge 的刚性与局部空隙,使分散在 branch migration staples 中的 toehold staples 仍能有效捕获 invader。这里的关键不是多几个 sticky ends,而是证明 tile-scale edge alignment 能降低分散 toehold 的熵惩罚;这是区别于 strand displacement 直觉的地方。
3. 避免 self-occluding edge:解决的是设计空间中“看似可用、实际拖慢/污染”的反应。ExGx 这类边缘会通过同一 tile edge 上的序列互补导致 invader self-dimerization 或 spurious branch migration。这个约束本质上是对可组合设计空间的剪枝,比单纯报告更多 reaction identities 更重要。
4. multitile invader assembly rules:解决的是复杂 invader 的产率问题。active external edge 必须足够强以触发 displacement,但它也会和内部 edge 发生竞争。作者通过削弱内部边缘而不是削弱 external active edge,使 undesired assembly 的 melting window 下移,同时保持 desired invader 可形成。
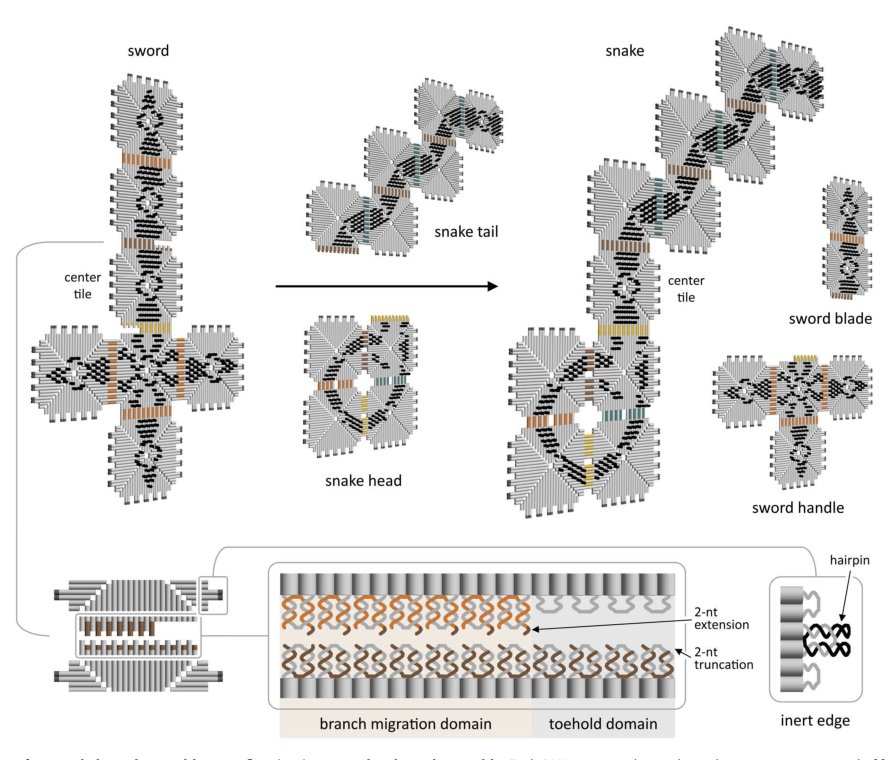
5. variable-size polymer / barrel opening:解决的是 invader 不一定固定尺寸的问题。tile ratio 控制 polymer 长度分布,embedded toehold 使 3D barrel 可被 tile displacement 打开成 2D chain。这个部分展示机制的几何外延,但更像 proof-of-mechanism,不是最终 modular robot 系统。
Key Insight / Why It Works
最核心 insight 是:在 tile 尺度上,specificity 可以从“binding equilibrium”迁移到“reaction pathway gating”。tile assembly 本身 specificity 不够,尤其在室温下容易错误粘;但 tile displacement 要求 invader 先找到开放 toehold,再经过 branch migration,错误结合即使发生也更难完成完整替换路径。这类似 strand displacement 提升 nucleic acid hybridization specificity 的机制,但放大到 origami tile level。
真正有效的部分我认为有三点。第一,mixed domain 和 discontinuous toehold 确实扩大了可编程空间,而且没有显著牺牲 kinetics;这是实质贡献。第二,self-occluding edge 的识别很重要,因为它指出 combinatorial design space 中大量“理论可编码”的反应在物理上不可用。第三,多 tile invader 的 internal/external edge competition 分析把问题从“能不能画出结构”拉回到“annealing pathway 是否会把 tile 用在错误位置”。这对后续系统设计比 sword-to-snake demo 本身更有价值。
相对而言,sword-to-snake 更像 integration demo;barrel opening 是有趣的几何扩展,但其增益来源不完全清晰,部分可能主要来自已有 tile polymer folding 与 capping 设计的 scaling / engineering。论文没有证明大规模并行网络,只证明了两个反应可以低串扰同时发生。所谓 robustness to temperature and concentration 目前也应理解为相对 assembly/disassembly 的机制优势,而不是已经完成系统级鲁棒性验证。
这不是 retrieval/data 覆盖类工作,而是 better inductive bias:通过 toehold-gated local replacement 把反应空间结构化。它的“推理”不是智能规划,而是物理反应路径编码。若未来声称可编程 molecular computation,真正瓶颈会从理论反应数转向错误模型、纯化、局部化、spurious occlusion 和路径级调度。
Relation To Prior Work
最接近的谱系有三条:DNA strand displacement、DNA origami tile assembly、modular self-reconfigurable robotics。论文的技术内核明显来自 strand displacement:toehold、branch migration、sequestered domains、kinetic control。但它不是简单把短链换成长链,而是要让整个 origami tile edge 作为 branch migration interface 工作,这引入了刚性、柔性、空间 occlusion 和 assembly pathway 等 strand-level 没有的约束。
相对于传统 DNA origami assembly / algorithmic self-assembly,差异在于它不是靠一次性 annealing 得到目标结构,也不是通过升温/降温改变 equilibrium,而是通过 invader-triggered replacement 在固定温度下改变结构。相对于 shape-complementary dynamic DNA devices,它更强调信息编码和反应级联潜力,而不是单个机械构件的几何锁合。
看似新的地方中,变长 polymer 控制、barrel folding、AFM 形状读出等不少是已有 DNA origami assembly 思想的重组;实质创新在于把这些结构作为 tile displacement invader 或 displacement substrate 来组织,并系统给出哪些 edge-domain 设计会扩大反应空间、哪些会造成 self-occlusion。它属于“DNA molecular robotics 的反应原语扩展”而不是“新型纳米结构制造”。
Dataset / Evaluation
评价是 wet-lab experimental validation,不是 dataset benchmark。覆盖范围包括最小 dimer 反应、复杂 sword/head/tail assembly、变长 polymer、3D barrel opening、两个并行 displacement reaction。实验工具主要是荧光动力学与 AFM imaging,足以支持机制层面的 proof-of-concept。
但 evaluation 对核心 claim 的支持有边界。它确实证明了:不同 domain 配置和 discontinuous toehold 可工作;self-occlusion 会损害 kinetics/yield;多 tile invader 需要考虑 assembly pathway;两个反应能在同一 test tube 中并行。然而它没有证明“hundreds of distinct reactions”可以共存,也没有证明复杂 artificial cell-like membrane 上的局部重配置。并行性只测试两个 invader,串扰也已观察到。AFM 计数受 sample prep、mica flattening、tip perturbation 和结构识别影响,尤其 barrel 部分需要间接推断;证据合理但不是完全闭环。
Limitation
最大限制是 scalability 被物理制造误差而不是理论编码空间主导。论文给出的 832 或 1104 这类 reaction number 是在 edge identity/toehold combinatorics 上算出来的,可用性还要经过 sequence-level cross-talk、shape occlusion、assembly purity、kinetic compatibility 筛选。实际系统里 active external edges 之间会互相 occlude,复杂 assembly 的不完整率也很高。
第二,invader 是 free-floating resource,不是被待重配置结构携带或局部化的组件。因此 sword-to-snake 更像环境中加入两个 reagent 后的化学转换,不是 electromechanical modular robot 意义上的 autonomous self-reconfiguration。作者也承认需要 localized tile displacement;否则 stochastic encounter 和 invader availability 会限制真正部署。
第三,机制依赖一个脆弱 trade-off:tile 需要足够柔性来允许 branch migration,又要足够刚性来保持功能分子定位和几何可预测性。更 flexible 的 wireframe 可能提升 displacement,但会损害 attached circuits/motors 的空间精度;更 rigid 的 tile 可能适合计算/运输,但替换变慢。文中未充分说明如何系统优化这个 trade-off。
第四,barrel-to-2D reconfiguration 的解释仍有间接性。AFM 上 barrel flattening/breaking 使结构统计带有样品制备偏差,荧光 evidence 支持 closed edges,但 intermediate geometry 文中未充分说明。最后,所谓 robust to temperature and tile concentration 目前更像相对优势,不应被解读为已解决真实复杂环境下的鲁棒部署。
Takeaway
- 1. Tile displacement 的价值在于把 DNA origami tile interaction 从 equilibrium assembly 改造成 path-gated reaction;这是向可编程 molecular robotics 迈进的关键抽象。
- 2. 扩大 design space 本身不够,必须同时给出“不可用设计”的负规则。
- self-occluding edge 和 internal/external edge competition 是这篇最值得迁移的工程-物理 insight。
- 3. 未来真正重要的方向不是再做更花的形状转换,而是 localized/catalytic invader、shape-complementary anti-occlusion、自动化 reaction compiler 和系统级 error model。
一句话总结
这篇论文把 DNA origami tile displacement 从小规模替换演示推进为一种可组合的模块重配置原语,真正贡献在于建立 edge-domain 设计空间与物理失效规则,而不是 sword-to-snake 这个具体形状转换。