精读笔记
Problem Setting
这篇论文实际在做 collective behavior 的“控制律识别”,而不是普通的鱼群行为描述。它试图回答:一个 juvenile zebrafish 在 schooling/following 时,究竟把看到的同伴信息怎样映射成自己的运动控制?
真正难点不在于观察到跟随,而在于自然交互里的因果结构不可辨识。两条鱼同时运动时,leader/follower 身份会切换,速度、位置、方向、视觉大小、遮挡、内部动机和环境边界全部混在一起。传统从轨迹反推 interaction rule 的方法容易得到一个看似合理的交互场,但很难判断它是鱼真实使用的输入输出映射,还是数据分布下的统计投影。
关键矛盾是:群体行为的宏观模型需要足够简单才能 scale,但个体感知-运动机制又必须足够具体,才能解释动态闭环交互。作者选择从最小但可因果控制的 leader-follower pursuit 入手,把 schooling 的核心局部机制还原成一个可执行控制器。
Motivation
已有路线的不足主要有两类。第一,传统生物群体模型多是从自然轨迹做反演,缺少干预,因此只能说“某种规则能拟合数据”,很难说“动物确实使用了这种感知变量”。第二,工程控制器如 MPC/PD 虽然形式清楚,但通常不是从动物的真实感知-行为闭环中反推出的,和生物机制之间缺少因果桥梁。
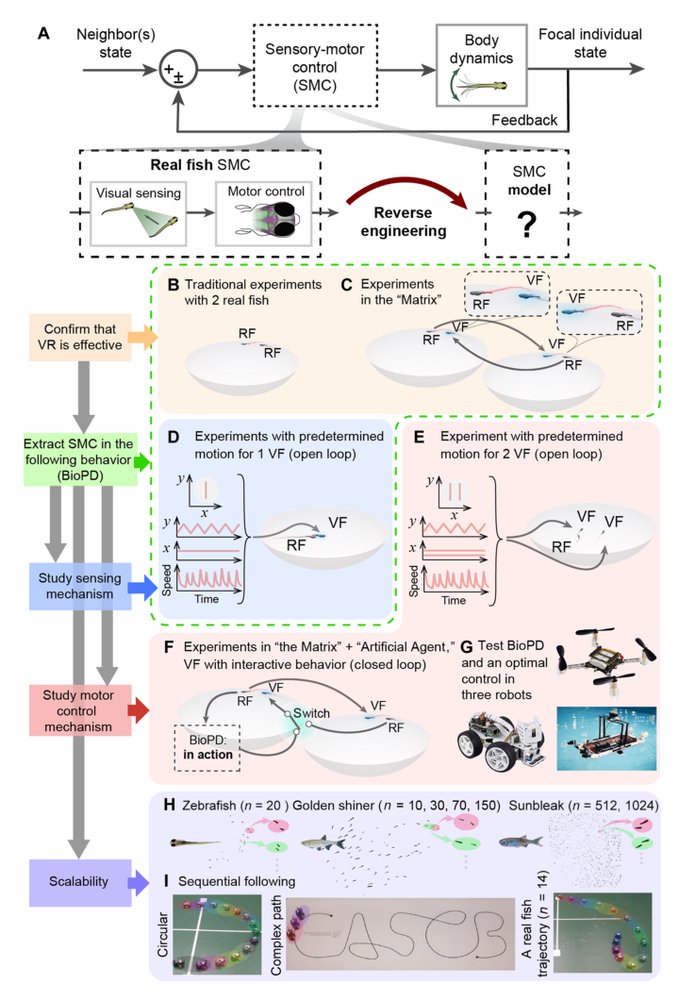
作者的核心观察是:VR 可以把社会交互变成类似神经科学 dynamic clamp 的问题。只要虚拟同伴被真实鱼行为上接受,就可以系统操纵虚拟同伴的速度结构、可见性、位置更新率和多目标布局,从而拆开“瞬时速度 cue”“平均速度 cue”“位置 cue”“生物运动模式 cue”等在真实世界中高度共变的变量。
因此这篇论文缺口意识很明确:不是再提出一个 social force model,而是建立一个能做 causal reverse engineering 的实验-模型闭环,用虚拟 conspecific 直接测试候选控制律是否能在真实动物面前成立。
Core Idea
核心思想是把 zebrafish schooling 中的 social response 压缩成一个 egocentric pursuit controller:鱼主要根据同伴在自身/leader 局部坐标中的相对位置,以及由位置随时间变化得到的低通速度信息,分别控制 lateral 与 forward 运动。BioPD 不是复杂的多智能体策略,也不是显式 alignment-rule 的堆叠,而是一个带空间衰减的 PD-like controller。
这个建模方式改变了问题表述:从“群体如何保持队形”变成“个体如何追踪一个或多个视觉目标”。它引入的 inductive bias 很强:局部、低维、位置优先、平均速度而非瞬时速度、有限交互范围。正因为这个 bias 强,模型才可能在小数据、少参数、跨速度、跨平台时表现稳定。
和 prior 的本质区别在于,作者不是先验写一个吸引/排斥/对齐规则再做拟合,而是用 VR 把输入空间拆开,反推一个可闭环执行的控制律,并让真实鱼与该控制律控制的虚拟体交互。这里最重要的新增信息不是 PD 形式本身,而是证明在这个生物系统中,位置驱动的低通 pursuit 足以解释一组关键 social response。
Method
1. VR/Matrix 作为因果接口:它解决自然交互中输入不可控的问题。通过让真实鱼与虚拟鱼或另一个真实鱼的实时投影交互,作者能固定或替换其中一方的控制策略。核心变化是把群体行为实验从被动观测变成闭环干预。
2. 单 leader open-loop 反推控制律:虚拟鱼沿可控轨迹运动,真实鱼作为 follower。作者把 follower 的运动分解到 leader-centric 坐标系里的 lateral 和 forward 分量,观察速度输出如何依赖相对位置与 leader 平均速度。这一步解决的是从行为场到控制律的识别问题。
3. BioPD 形式:控制律是 proportional-derivative-like,但乘以 Gaussian derivative 型的空间权重,使反应在约 1 个特定距离尺度附近最强,远处衰减。这解决了传统线性 PD 无限作用范围不符合社会交互的问题,也让控制律具有生物上合理的 cutoff。
4. 感知假设验证:通过 constant speed vs burst-glide、不同 update frequency、周期性不可见、不可见期间位置/速度不一致等干预,作者测试鱼到底响应什么。结论是瞬时速度和生物运动细节不是主要输入,鱼更像是利用位置序列积分出平均运动信息。
5. 多目标与闭环验证:双 leader 实验和 Matrix Turing test 用来检查 BioPD 是否只是单 leader 拟合,还是能进入更真实的 reciprocal interaction。双目标下的 bifurcation 是很关键的外推测试,因为它要求模型不仅复现均值距离,还要复现决策几何。
Key Insight / Why It Works
最关键的 insight 是:schooling 的一个核心局部机制可能不是 alignment-dominated,而是 pursuit-dominated。鱼不需要显式估计同伴瞬时速度,也不需要解析复杂生物运动模式;只要有一个相对位置表征,并在约 0.2 s 量级上整合位置变化,就可以实现稳定跟随、速度匹配和多目标选择。
BioPD 有效的主要原因不是 PD 本身新,而是它把正确的变量选对了:relative position in egocentric/leader-centric coordinates。这个 representation alignment 很重要。若用全局坐标或直接速度 cue,模型会更依赖场景和噪声;用相对位置则自然适配 leader 转向、速度变化和视觉遮挡中的短时缺失。
另一个核心原因是低通/平均速度机制。鱼对 burst-glide 与 constant-speed 反应类似,对低于约 5 Hz 的更新才明显退化,这说明 fine-scale locomotor signature 不是 schooling 控制的主变量。这个结果削弱了“生物运动模式是主要 social cue”的解释,支持“位置记忆 + 平均运动估计”的控制视角。
空间 cutoff 是 BioPD 比普通工程 PD 更像生物交互的地方。没有 cutoff,PD 会变成无限范围追踪器;加入一阶 Gaussian derivative 后,模型自然产生一个最佳作用距离,并在远距离减弱。这解释了为什么 lateral/forward response 都有一个峰值距离,而不是随误差单调增加。
哪些部分可能只是辅助?机器人实验主要是展示 BioPD 作为 kinematic pursuit heuristic 的工程可用性,不能反过来强力证明生物机制。大群体跨物种分析也更像一致性证据:说明局部 leader-following signature 在更大群体中存在,但未充分证明同一参数、同一控制律在高密度多邻居交互中严格成立。
如果做技术归因,这篇的贡献更接近 better inductive bias + causal experimental identification,而不是 scaling、RL、test-time compute 或复杂优化。它的强处在于把可解释控制律嵌入真实闭环生物交互并通过干预排除若干替代解释。
Relation To Prior Work
它最接近三条谱系:动物群体交互规则反演、bio-inspired control、dynamic clamp/VR 行为干预。和经典 attraction-alignment-repulsion 或 social force 模型相比,它不是从宏观队形倒推局部势场,而是直接识别感知输入到运动输出的控制律。和已有 PD-like biological controller 工作相比,PD 形式并不新,新增点在于通过 VR 进行因果拆解,并把同一个控制律放回真实社交闭环中测试。
与 Laan/de Polavieja 一类从双鱼轨迹中寻找 optimal control signature 的工作相比,这篇的本质进步是可干预性:它能让 leader 的速度结构、可见性和更新率独立变化,从而验证输入变量,而不只是从自然共变数据里拟合。与物理机器人鱼/replica 研究相比,VR 的优势是能更精细地解耦位置和速度感知;实体 replica 很难做到突然不可见、位置跳变、视觉更新率操纵且不引入其他机械/水动力 confound。
看似新的地方中,机器人跨平台应用更多是已有 pursuit control 思想的生物参数化重组。实质创新是“dynamic social clamp + experimentally derived BioPD + embodied Turing test”这一组合:它让一个低维控制律不仅解释数据,还成为交互对象的一部分。
Dataset / Evaluation
评估覆盖面相当完整,尤其对核心 claim 支持较强。它包括 VR 系统与真实双鱼交互的行为等价验证、单虚拟 leader open-loop 识别、感知变量消融、动态真实 leader 轨迹模拟、Matrix 中真实-虚拟闭环替换、双虚拟 leader 多目标预测、跨物种大群体轨迹分析,以及三类机器人真机追踪。
真正支持核心生物 claim 的是前五类:VR 等价性、open-loop 反推、感知消融、动态 leader 验证和 Turing test。这些实验共同证明 BioPD 不是简单 overfit 到一条固定轨迹。双 leader bifurcation 是很强的预测性测试,因为它要求模型产生定性结构变化,而不只是匹配平均距离。
跨物种/大群体和机器人部分支持的是 generality claim,但力度不同。大群体数据表明类似 leader-following signature 可在更大规模出现,但由于不是同样强干预,因果性较弱。机器人实验有真实世界真机价值,但任务相对受控,更多证明 BioPD 足够 robust 和 cheap,而不是证明它在复杂 autonomous navigation 中优于优化控制。
总体看,evaluation 对“BioPD 是 juvenile zebrafish social pursuit 的有效算法级模型”支持充分;对“这是通用群体控制律”或“工程上接近最优”支持较弱,需要更复杂环境、约束、扰动和多邻居密度下验证。
Limitation
第一,模型成立依赖视觉主导和同平面交互。作者明确指出 juvenile zebrafish 在该阶段主要依赖视觉,且 Matrix 中只有处于同一 x-y plane 时交互强。这个前提限制很大:三维 schooling、成鱼、多感官整合、水动力 cue 更强的条件下,BioPD 可能只是子机制。
第二,感知前端未解释。文中证明鱼行为上像是在使用 positional information,但鱼如何从视觉场中估计同伴位置、距离、朝向,文中未充分说明。这个问题不是细节,而是控制律能否真正落到神经/感知机制上的关键缺口。
第三,多邻居扩展仍不充分。双 leader 实验显示模型能预测 averaging-to-choice bifurcation,但真实大群体中存在遮挡、视野限制、多个局部 leader、身份切换和密度依赖。BioPD 是否以简单叠加形式扩展,还是需要 attention/winner-take-all/visibility weighting,文中未完全回答。
第四,参数泛化的边界不清。Kp、Kd、rx、ry 是从 juvenile zebrafish 和特定 VR setup 估计的。机器人实验中通过 body-size scaling 使用相同参数,这很漂亮,但增益来源不清:可能主要来自任务尺度和速度范围温和,而非参数具有普适最优性。
第五,机器人应用可能夸大了“接近最优”的含义。MPC 在每个平台上优化,BioPD 用生物参数仍接近其控制能耗,这很有趣;但测试是追踪平滑虚拟 leader,未包含复杂障碍、碰撞约束、执行器饱和、安全距离、多目标冲突和延迟。这里的性能可能主要来自 scaling / task simplicity,而不是 BioPD 具备规划能力。
第六,BioPD 是算法级充分模型,不是唯一模型。存在其他低维控制律也可能复现实验现象,尤其在相同输入变量和类似 cutoff 下。论文的因果干预排除了若干替代解释,但没有证明唯一性。
Takeaway
- 1. 这篇真正推动的是 collective behavior 的因果 reverse engineering:用 VR 把社会交互变量拆开,再把候选控制律放回真实闭环里检验。
- 这比单纯轨迹拟合更接近机制识别。
- 2. 对 schooling 来说,一个值得迁移的观点是:局部 pursuit primitive 可能比传统 alignment rule 更基础;alignment 可能是追踪多个移动目标后涌现出的宏观现象,而不是个体显式计算的目标。
- 3. 低通的位置记忆是关键设计原则。
一句话总结
这篇论文把斑马鱼 schooling 的局部社会响应从统计交互规则推进到可因果验证的低维 pursuit control law,核心贡献是用 VR/dynamic social clamp 证明位置驱动、低通、带空间 cutoff 的 BioPD 足以解释并嵌入真实闭环跟随行为。