精读笔记
Problem Setting
【Artificial embodied circuits uncover neural architectures of vertebrate visuomotor behaviors,Science Robotics / 2025】
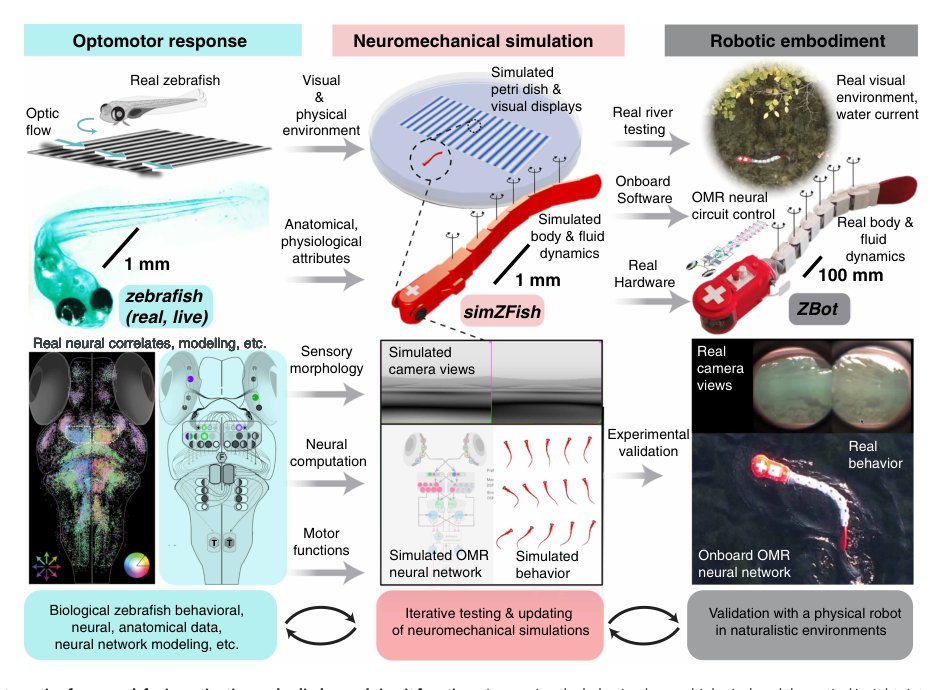
这篇论文真正处理的是 embodied neuroscience 里的一个硬问题:神经回路的功能不能只从神经活动或刺激-响应相关性中读出,必须放在身体和环境闭环里验证。具体到 larval zebrafish OMR,已有工作已经知道许多相关脑区和响应类型,但仍不清楚这些响应类型为何如此组织、哪些连接是功能必需、哪些是实验刺激空间下的拟合产物。
关键矛盾是:真实动物中最想操纵的变量——透镜、视野、retina-pretectum 连接、身体水动力、视觉反馈——几乎不能独立操纵;而纯计算模型又通常缺少真实感官输入和运动反馈,因此只能解释神经活动,不能解释行为稳定性。作者把问题重写为:一个基于真实神经数据的人工回路,放入逼真的鱼体和环境后,是否能复现 OMR,并通过反事实操纵解释真实神经架构。
Motivation
已有路线的问题不是数据少,而是数据和闭环功能之间断裂。全脑成像可以告诉我们哪些 neuron 对某类 optic flow 响应;行为实验可以告诉我们鱼如何转向;但二者之间缺一个可运行的、从像素到肌肉的中间对象。没有这个对象,就很难判断某个 response type 是控制变量、感觉副产物,还是实验刺激设计诱导出来的分类。
作者的核心观察是:斑马鱼的视觉运动计算高度依赖 embodied geometry。侧向眼、鱼眼透镜、下视野纹理、Snell’s window、底部投影条纹、局部 DSGC motion detector 都会改变进入 pretectum 的有效光流。因此,很多“神经选择性”可能不是纯粹由内部回路决定,而是由身体-环境共同筛选出来。缺口正是一个能系统改变这些物理和连接变量的实验平台。
Core Idea
核心思想是把 OMR 回路做成一个人工 embodied circuit,而不是一个神经响应拟合模型。simZFish 从模拟视网膜像素开始,经 DSGC/ePT/lPT/nMLF/aHB/CPG 等抽象回路,到尾部运动,再通过水动力和视觉反馈回到下一帧感官输入。这样,网络必须在闭环中稳定自身,而不是只在离线 stimulus 上拟合 calcium trace。
本质区别在于评价对象变了:prior 多数是在“刺激 → 神经活动”或“刺激 → 行为”的两个投影上做建模;本文把二者合成一个可执行的因果系统,并用物理身体作为正则化。这个 inductive bias 很强:只有那些同时兼容 retinal optic flow、局部运动检测、运动控制和反馈稳定性的神经结构,才会表现好。它不是单纯 scaling,也不是更大数据拟合,而是把缺失的 latent structure——身体和环境几何——显式放回模型。
Method
方法上值得保留的机制只有几类。
第一,端到端闭环神经-机械仿真。它解决的是以往 OMR 模型不能产生真实运动反馈的问题。通过让人工神经活动直接驱动 bout initiation、turn bias 和尾部 CPG,模型的中间表征必须对行为有可检验后果。
第二,感官几何反事实操纵。透镜焦距、视野范围、retina-pretectum 连接象限不是作为生物细节加入,而是用于回答“视觉输入的物理投影如何塑造神经架构”。核心变化是把 receptive field 偏置从描述事实变成可解释约束:下后方视野在该 embodied setup 中提供更一致、更少冲突的 forward/medial optic-flow signal。
第三,模型失败驱动实验迭代。simZFish1.0 在未见过的 parallel forward/backward 组合刺激上预测错误转向,作者没有把它当作 bad fit,而是用它定位缺失神经计算,再通过真实鱼行为和双光子成像发现 forward/backward selective subtypes,更新为 simZFish2.0。这是论文中最像“模型产生新神经假设”的部分。
第四,物理机器人验证。ZBot 的意义不是严格缩放幼鱼,而是测试同一视觉-运动算法在真实视觉噪声、湍流、浑浊和非理想控制下是否仍提供稳定增益。
Key Insight / Why It Works
最重要的 insight 是:OMR 回路中的某些结构看似是 neural specificity,实际上可以由 embodied optic-flow statistics 推导出来。特别是下后方视野的优势,不只是“那里有更多纹理”这么简单,而是侧向眼看到底部平移刺激时,局部光流会因透视畸变产生旋转分量;如果全视野 DSGC 都接入,下前方与下后方的局部运动信号会互相抵消或混淆。限制到下后方视野反而把 forward/medial motion 变成下游回路更容易利用的信号。这是典型的 better inductive bias,而非 scaling。
方法有效的第二个原因是 closed-loop evaluation 暴露了开环模型看不到的错误。simZFish1.0 能拟合原始 OMR 条件,但在 conflicting forward/backward stimulus 上出现真实鱼没有的强转向。这说明原先 response classes 覆盖不足;新增 forward/backward subtypes 后改善,表明模型失败确实对应了缺失的 latent neural structure,而不是单纯调参。
第三,ZBot 结果说明该回路抓住了一个低维控制规律:由水流拖拽引起的 optic flow 足以提供 upstream orientation cue。这里的“rheotaxis”更像一个 emergent stabilizing controller,而不是复杂导航或规划。不能把它解读成高级空间推理;它本质是视觉反馈控制。
哪些可能只是辅助?Webots 高保真、机器人外形、CPG 细节和大量参数调节都重要,但更偏 engineering。论文真正的贡献不是仿得多像,而是用 embodied counterfactuals 解释 neural architecture,并让模型失败产生新实验。增益来源中仍有不清楚的部分:simZFish 初始化于已有 best-fit 连接权重,后续又经过最小调参;文中未充分说明不同参数化是否会得到同样的结构结论。
Relation To Prior Work
这篇处在 neuromechanical simulation、bio-robotics 和 zebrafish whole-brain circuit modeling 的交叉谱系上。最接近的不是一般机器人控制,而是 NeuroMechFly、virtual rodent、lamprey/salamander CPG robot、以及 Naumann 2016 的 zebrafish OMR circuit model。
相对 Naumann 2016 等工作,真正新增的是 embodied closure:原先的功能回路主要解释视觉刺激下的神经响应和行为相关性;本文让同一类回路在物理环境中产生行为,并允许操纵视觉形态和连接。相对 NeuroMechFly/virtual rodent,本文的创新不在大规模学习或运动控制复杂度,而在把具体的 vertebrate visuomotor circuit 与真实神经成像迭代结合。
看似新的地方中,机器人验证、CPG 控制、光流稳定控制本身都不是新思想;实质创新是把这些工具组织成一个“从生物数据 → embodied model → 反事实预测 → 新神经实验 → 真实机器人”的闭环科学工作流。它更像一种研究范式贡献,而不是某个单点算法突破。
Dataset / Evaluation
evaluation 覆盖了四类证据:实验室 OMR 行为复现、神经活动对齐、虚拟河流 rheotaxis、真实机器人河流测试。这个组合基本支撑作者的核心 claim:embodiment 对解释神经架构有实际作用,而且人工 OMR 回路可以在未完全控制的真实环境中产生稳定增益。
但需要注意,任务覆盖仍集中在 OMR/position stabilization 这一类低维视觉反馈行为。所谓跨场景泛化主要是从 drifting gratings 到虚拟/真实河流视觉流,而不是跨任务泛化。真实机器人验证有价值,因为它引入真实视觉和流体复杂性;但 ZBot 与幼鱼尺度差异太大,水动力 regime 不同,因此它验证的是控制算法的现实可用性,不是严格验证生物力学等价。
benchmark 是否验证 claim?对“视觉 OMR 回路足以辅助 rheotaxis”支持较强;对“真实神经架构由 embodiment 决定”支持中等偏强,但仍依赖模型假设。因为可能存在其他回路结构也能达成同等行为,只是与当前已知神经数据不一致。文中承认 lower anterior alternative 可能可行,但需要不同 downstream circuit;这说明 optimality claim 应理解为在给定已知 zebrafish circuit family 内的最优,而非全局最优。
Limitation
最大限制是可解释性来自强先验,而不是自动发现。网络结构、神经类别、连接方向、bout gate、CPG、turn determinator 都带有大量已有生物知识和工程抽象;模型能工作并不意味着这些抽象层级就是生物真实机制。尤其 motor system 被高度简化,很多 downstream spinal dynamics 和 sensory reafference 只是功能替代。
第二,泛化上限有限。OMR/rheotaxis 是非常适合 optical-flow feedback 的任务,控制变量低维且环境 cue 强;在 prey capture、escape、multi-sensory arbitration 或 internal-state-dependent behavior 中,当前框架是否仍能扩展并不清楚。没有 motor adaptation/gain control 时,simZFish 和 ZBot 在较快流速下都会失败,说明 controller 仍是固定增益反射系统。
第三,增益归因不完全干净。simZFish1.0 到 2.0 的改进来自新增 neural subtypes,但也可能部分来自更好的 stimulus coverage 和手动结构修补。文中未充分说明是否存在系统 ablation 来区分 subtype identity、connectivity weight、threshold/bout policy 各自贡献。
第四,ZBot 真实河流实验样本量小、环境不可完全控制,且 blinded OMR 条件下随机游泳本身也显著帮助稳定位置。OMR 的额外增益是真实的,但不是压倒性的;它说明视觉回路有用,不说明它足以完整解决自然 rheotaxis。
Takeaway
- 1. 对 sensorimotor neuroscience 来说,这篇最值得迁移的是“模型失败作为实验设计器”:先让 embodied model 在新刺激上犯错,再用真实神经数据定位缺失 computation。
- 2. 神经架构解释不能只看回路内部;感官形态和环境统计可能直接决定哪些连接是有效的。
- 这里的 lower-posterior visual field 是一个很好的例子。
- 3. 对机器人方向的启发是:低维生物反射回路在真实环境中可能比端到端学习控制器更可解释、更轻量,尤其适合稳定、对齐、姿态保持这类任务。
一句话总结
这篇论文把斑马鱼 OMR 从离线神经响应模型推进到可操纵的 embodied closed-loop circuit,用身体和环境几何反推神经架构约束,是一篇范式型的 neuromechanical neuroscience 工作,而不是单纯仿生机器人或仿真工程。