精读笔记
Problem Setting
这篇论文实际在解决一个“传感器为什么长成这样”的控制论归因问题:苍蝇 LPTC 的空间调谐是否只是 optic-flow matched filtering,还是反映了飞行 plant 的输入-状态-输出能量结构。
真正困难点在于跨层映射。LPTC response field 是视觉神经生理对象;飞行自然模态、可控性、扰动敏感性是动力学对象;控制输入又不是飞机舵面那样清楚定义,而是变形翼拍动的高维周期运动。过去工作通常只覆盖其中一端:要么解释 LPTC 对光流的匹配,要么建昆虫刚体飞行动力学,要么做 neuromechanical simulation,但缺少能抽象出控制论原则的解析模型。
关键矛盾是:工程直觉会把传感器放在最大化状态估计的位置;昆虫可能根本不需要估计完整状态,而是直接检测那些最可能被自身动作或阵风激发、且对闭环稳定最有价值的低维运动组合。
Motivation
已有路线不够的原因是它们没有回答“matched filter matched to what”。LPTC 像 matched filters 这一点并不新;新问题是这些 matched directions 是否由视觉生态决定、由自然模态决定、由可观测性决定,还是由控制输入和扰动共同决定。
作者的关键观察是:资源受限的昆虫不太可能采用现代飞控式的 dense sensing + recursive state estimation。更合理的设计是把物理系统中最有控制价值的状态方向预先编码进传感器形态和神经连接中,让 sensory front-end 本身完成一部分模型降维和控制相关特征提取。
因此缺口不是“缺一个更精细的昆虫模型”,而是缺一个能同时比较 controllability、observability、disturbance sensitivity 及其 joint objective 的统一框架。
Core Idea
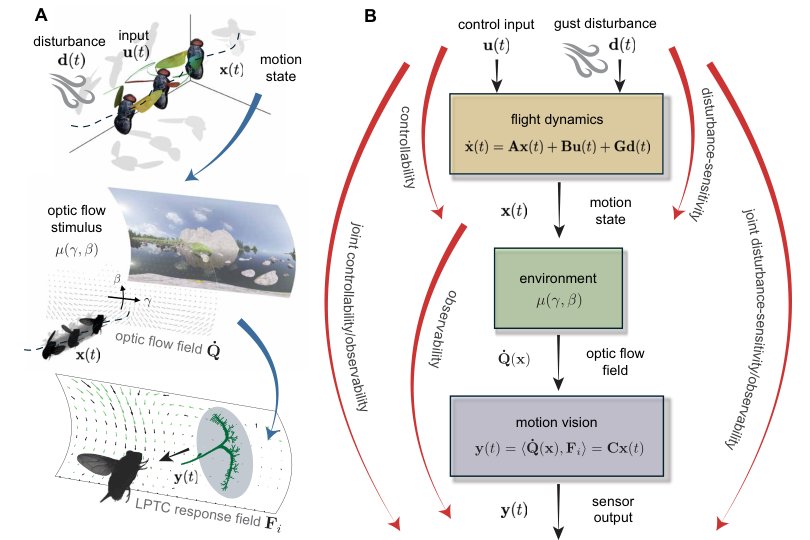
核心思想是把 blowfly motion vision 看成一个 plant-aware output map:控制输入 u 和阵风扰动 d 经飞行动力学 A/B/G 激发状态 x,状态 x 通过光流和 LPTC response fields 形成传感输出 y=Cx。LPTC 的 preferred self-motion directions 就是 C 的行向量;问题变成这些行向量是否对齐到系统最重要的能量传递方向。
理论直觉很清楚:如果某个状态方向很容易被执行器驱动或被扰动激发,但传感器看不到,那闭环控制会低效;如果某个方向传感器很敏感但几乎不会被输入/扰动激发,那它对飞行稳定的能量价值有限。最有用的方向是两者兼具,即 controllable/observable 或 disturbance-sensitive/observable。HSV 正好度量这种输入到输出经状态传递的能量能力。
本质区别在于它没有把 sensing 作为 state estimation 的后处理问题,而是把 sensing、actuation、disturbance 和 body dynamics 作为共同设计对象。这是一个 embodied control 的解释,而不是一个视觉编码解释。
Method
1. 端到端状态空间化。作者把飞行线性化为 xdot=Ax+Bu+Gd, y=Cx。A/G 来自刚体动力学和 CFD 稳定导数,B 来自自由飞行翼运动的功能主成分,C 来自 LPTC response fields 与自运动光流偏导的内积。必要性在于只有这样才能把“神经元偏好的光流”转成“状态空间中的方向”。
2. 用 Gramians 定义三类重要方向。Controllability Gramian 给出最省控制能量可达的状态方向;disturbance-sensitivity Gramian 给出最容易被阵风激发的方向;observability Gramian 给出最容易释放到传感输出的方向。它解决的是 mode-sensing hypothesis 过去过于定性的问题。
3. 用 balancing / HSV 做坐标无关的 joint test。单看某个 Gramian 会依赖状态变量如何缩放和表示;HSV 作为相似变换不变量,更适合支持“系统层面能量吞吐最大化”的 claim。作者用随机镜像 LPTC direction sets 构造 null distribution,测试真实 LPTC 是否处在高 HSV 区域。
4. 分细胞亚群解释功能分化。VS/V 更贴近稳定控制相关方向,HS/H 更偏向导航/稳态运动相关方向。这里不是主贡献,但增加了生物解释的可信度。
Key Insight / Why It Works
最重要的 insight 是:昆虫视觉输出层可能不是为“重构世界/重构完整状态”服务,而是为“最大化控制相关信号能量吞吐”服务。换句话说,LPTC 做的不是通用感知,而是一个与 plant dynamics 共同调谐的低维投影。
方法有效的核心在 HSV 而不是在单独的 Gramian 对齐。单独说 VS 细胞对齐最可控方向或最扰动敏感方向仍可能受坐标选择、尺度归一化和模型参数影响;但 joint controllability/observability 的 HSV 解释更强,因为它对应输入到输出的 Hankel 能量通道,并且能连接到闭环控制中的最小执行器努力和鲁棒性边界。这使得论文的 claim 从“像自然模态”升级为“像优化开环 plant 以提升潜在闭环性能”。
最可能是核心贡献的部分:把 LPTC preferred directions、飞行动力学、控制输入、阵风扰动放进同一个 balanced realization 语言里,并证明真实传感器方向在随机传感器空间中接近能量吞吐上界。这个贡献是 representation alignment,不是 scaling。
可能只是辅助或 engineering 的部分:高精度视频、x-ray inertia、RANS、V-cell 新电生理数据提高了模型可信度,但不是概念突破本身。它们服务于让 A/B/G/C 足够可信,避免控制论分析变成空中楼阁。增益来源主要来自建模闭环链条的完整性,而非某个单一数据模块。
需要警惕的是,B 的定义可能显著影响 controllability directions。功能 PC 是从自然飞行数据中学到的低维控制基,天然包含行为分布先验;因此“LPTC 对齐可控方向”部分可能同时反映了真实控制能力和采样到的 wingbeat variation。这里不是 benchmark leakage,但有 implicit behavioral prior。文中没有完全拆清楚该 prior 对结果的贡献。
Relation To Prior Work
最接近的谱系有三条。第一是 LPTC matched-filter / optic-flow coding 工作,它们说明 LPTC 对特定自运动光流调谐,但没有解释这些自运动为什么动态重要。第二是 insect flight dynamics / stability 模型,它们分析自然模态和不稳定性,但通常没有 sensory output equation。第三是 sensor/actuator placement、balanced truncation、Hankel norm 相关控制理论,它们提供了 joint controllability-observability 的语言。
这篇真正新增的信息是把这三条线合在一起,并把 mode-sensing hypothesis 形式化为 HSV 最大化。看似新的“传感器检测自然模态”其实不是新概念;实质创新是指出自然选择可能优化的不是 natural eigenmodes 本身,也不是 observability alone,而是 input/disturbance-to-output signal energy transfer。
与传统工程传感器布置的本质差异在于目标函数不同。工程上常见目标是让状态估计最好;这里的目标是让由执行器和扰动实际激发的状态方向在传感端最强可见。它更像 early-stage co-design / embodied sensing,而不是 late-stage sensor placement。
Dataset / Evaluation
评价覆盖了真实生物数据和模型仿真两端:LPTC response fields 来自新旧电生理数据,自由飞行翼运动来自高速视频,惯量来自 synchrotron micro-CT,气动导数来自 RANS。严格说这不是传统 dataset/benchmark 论文,而是模型归因论文。
是否支持核心 claim?总体上支持。最关键的评价不是某个飞行性能指标,而是真实 LPTC 构成的 C 是否比随机 C 产生更高的 HSV。作者还测试了 unilateral/bilateral 输出、不同环境 nearness 设定,说明结论不完全依赖某个任意组合方式或 2m cube 场景。
但 evaluation 没有直接验证闭环行为收益。论文证明了 open-loop plant 的能量吞吐优势,并引用理论说明这会影响最坏控制能量和鲁棒性;但没有在真实苍蝇或机器人上展示使用这种传感结构后闭环控制更省能、更稳或更快。核心 claim 是控制论解释层面的强证据,不是 deployment 级验证。
Limitation
1. 线性化和平衡飞行前提很强。分析围绕 level forward flight 的小扰动 LTI 模型展开;而真实昆虫飞行包含 saccade、非线性气动、状态依赖视觉增益和行为切换。泛化到强机动并非自动成立。
2. 控制输入定义有建模自由度。用 wing kinematics PCs 表示控制输入很合理,但它把观察到的运动统计和潜在可控子空间混在一起。前四个 PC 加频率是否是生物真实执行器空间,文中未充分说明。结论可能部分依赖这个 control basis。
3. C 的构造依赖视觉环境和头部姿态假设。作者做了 nearness 扰动鲁棒性,但真实环境的深度分布、遮挡、纹理、头部补偿和多模态融合都会改变 optic-flow-to-output map。这里的鲁棒性测试覆盖有限。
4. 因果性不足。论文证明“现有调谐与 HSV 最大化一致”,但没有证明“HSV 最大化导致该调谐”或“破坏这种调谐会降低真实飞行性能”。这类生物最优性论证天然有多解释空间。
5. 下游控制器缺失。HSV 是 open-loop 指标;论文合理地用控制理论连接到潜在 closed-loop performance,但实际神经控制器 K 未建模。若下游读出不是 balanced-coordinate friendly,能量吞吐优势可能无法完全兑现。
6. 可能主要来自 representation alignment,而非完整 sensorimotor theory。它解释了视觉输出层为什么对齐到某些动态方向,但还没有解释多模态融合、肌肉冗余、行为目标切换和学习/适应如何共同参与。
Takeaway
- 1. 最值得迁移的不是“仿苍蝇光流”,而是用 Hankel/Gramian 目标做 sensor-actuator-dynamics co-design:传感器前端应对齐到输入和扰动实际会激发且输出可读的状态方向。
- 2. 对微型机器人尤其重要:在算力、功耗、通信带宽受限时,把控制相关投影前移到传感硬件/前端,比 dense sensing 后接 Kalman filter 更有架构优势。
- 3. 这篇把 mode-sensing hypothesis 从漂亮直觉变成了可检验的系统理论命题。
- 未来真正值得做的是闭环验证:固定 plant,比较 observability-only、HSV-optimized、random sensors 在能耗、鲁棒性、延迟下的真实控制差异。
一句话总结
这篇论文的真正贡献是把苍蝇运动视觉从 optic-flow matched filtering 重新解释为一种 plant-aware、Hankel-energy-optimized 的 embodied sensing 设计原则,推动了仿生飞控从“模仿传感器形态”走向“联合优化传感-执行-动力学信息流”。